Biologia della
memoria:
dalla registrazione
dell'informazione al ri-consolidamento
L’engramma
1903: Rickard Semon Die Mneme (Mnemosine, dea della
memoria).
Semon intendeva unificare il campo dell'eredità con quello
della memoria: entrambi sarebbero dipesi da una memoria responsabile
della trasmissione dell'informazione di generazione in generazione
(la memoria genetica) oppure della trasmissione dei ricordi.
Tre diversi stadi: la “scrittura” nelle trame nervose
(engrafia), la traccia mnemonica o “engramma” e la
rievocazione o “ecforia”.
Marcel Proust: esiste un rapporto di
causa ed effetto tra stimolo e engramma?
“E come in quel gioco, che piace ai giapponesi, di buttare
in una ciotola di porcellana piena d'acqua dei pezzettini di carta a
tutta prima indefinibili che, non appena immersi, si stirano,
assumono contorni e colori [...] così, ora, tutti i
fiori del nostro giardino e quelli del parco di casa Swann, e le
ninfee della Vivonne, e la brava gente del villaggio [...] e
tutta Combray e la campagna circostante, tutto questo che sta
prendendo forma e solidità è uscito, città e
giardini, dalla mia tazza di tè.”
Karl Lashley (1890-1958): The search for
the engram
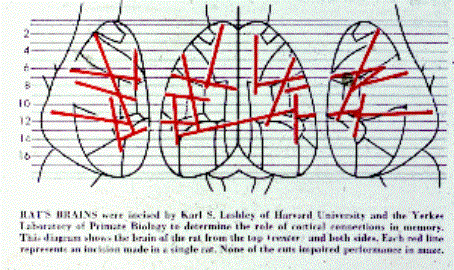
Il principio di massa d’azione e di
equipotenzialità
Per Lashley, olista convinto, era il cervello -o megli la
corteccia- nel suo insieme a codificare le memorie: praticando
lesioni in diverse aree corticali dimostrò che gli effetti non
dipendevano dalla sede ma dall'estensione della lesione.
Il test, le lesioni e le
prestazioni dei ratti nell’esperimento di
Lashley
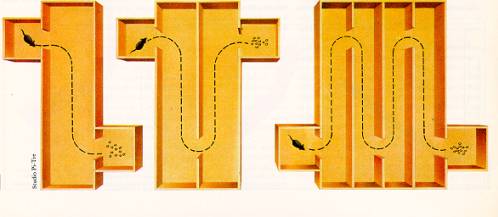
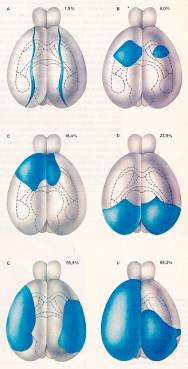
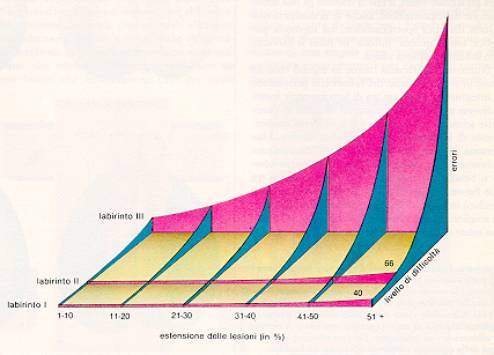
Il test messo a punto da Lashley era costituito da tre
labirinti di crescente complessità: il numero di errori
commessi dagli animali era proporzionale all'estensione della lesione
e alla difficoltà del compito: ciò avrebbe indicato che
il cervello era regolato da un principio di "azione di massa" e non
da specifiche localizzazioni delle funzioni. In realtà, il
test di Lashley misurava soprattutto variabili percettive
cenestesiche più che vere e proprie memorie.
Memoria a breve e a lungo
termine
H. Ebbinghaus e il metodo del
risparmio.
Nel 1873 aveva sostenuto una tesi di
filosofia su E. von Hartmann e l’inconscio che indicava, ben
prima di Sigmund Freud, una dimensione inconscia della memoria: il
metodo di Ebbinghaus dimostrava che nel ri-apprendimento si
verificava un risparmio e che ciò che veniva “trattenuto
in memoria” era indipendente dallo sforzo di rievocazione
cosciente.
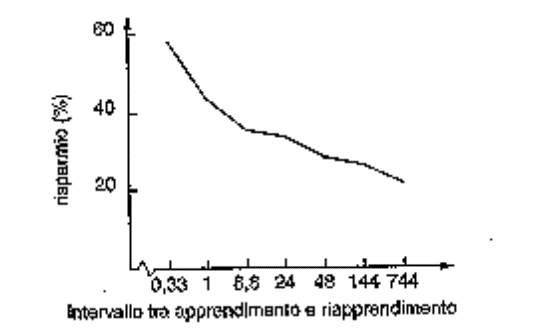
Ebbinghaus:
1. Il metodo del
risparmio
2. L’interferenza
La memoria verbale.
3 5 6
2 4 7 5
3 6 9 5 7
8 1 0 2 4 8
5 0 1 3 5 6 7
4 3 5 1 0 4 9 7
8 1 0 6 5 7 0 6 4
9 3 0 5 4 3 2 8 7 6
4 8 9 6 3 1 8 1 5 3 9
Dopo aver sentito leggere una serie di
numeri si è in grado di ripeterne 6-7.
Donald Hebb ( 1904-1985) e le
teorie “hebbiane”

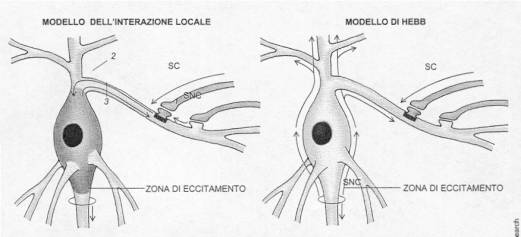
Nel modello dell’interazione locale
stimolo condizionato e non condizionato sono associati da sinapsi
adiacenti. L’eccitazione va e ritorna dal soma
(riverberazione).
Nel modello di Hebb i cambiamenti si
verificano se i due input (SC e SNC) coincidono.
Il consolidamento e
l’interferenza.
Il classico modello della memoria teorizzato da Hebb implica che
la memoria breve si trasformi in memoria a lungo termine grazie a un
processo di consolidamento.
La memoria a breve termine o “di
lavoro” comporta modifiche dell’attività
bioelettrica delle sinapsi.
Questa memoria è fragile e
diversi trattamenti fisici possono indurre amnesia.
L'elettroshock e il consolidamento della
memoria.
1949 C.P.
Duncan: Intervallo tra ECT e
apprendimento
J.Comp. Physiol. Psychol. 42, 3-44,
1949.
1960-1973 J.L.
McGaugh: Intensità ECT e
intrevallo tra ECT e apprendimento. One-trial
learning
Science 179, 1343, 1973.
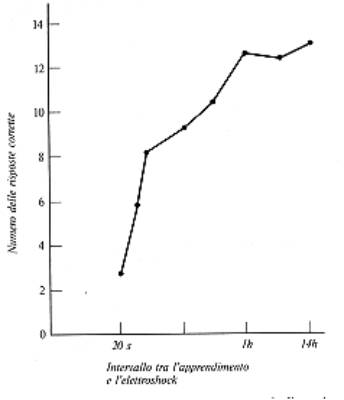
Duncan e McGaugh, attraverso una serie
di ricerche, hanno indicato che se una memoria a breve termine non
è stata ancora consolidata, alcuni trattamenti fisici come
l'elettroshock (ECS) hanno l'effetto di bloccare il consolidamento,
purchè l'ECS venga somministrato subito dopo
l'esperienza. Quando la memoria a breve termine è già
stata consolidata, l'ECS sarebbe inefficace: in realtà il
consolidamento non è stabile e negli ultimi anni si è
visto che la memoria è soggetta a continui rimaneggiamenti,
così da essere ri-consolidata.
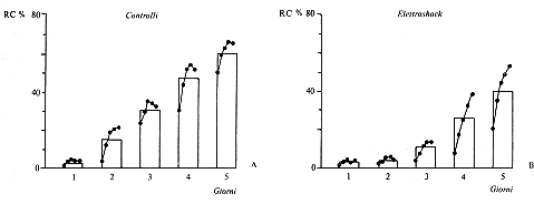
Il consolidamento della
memoria,
come ipotizzò Donald Hebb,
dipende dalla stabilizzazione delle sinapsi o dalla formazione di
nuove sinapsi. I trattamenti che interferiscono con la sintesi
proteica bloccano il consolidamento, cioè la formazione di
memorie “a lungo termine”.
1964 Agranoff e Klinger: Puromicina e
consolidamento in Carassius auratus.
Science 146, 952, 1964.

Il test utilizzato per studiare il consolidamento della memoria
nel pesce Carassius auratus, il comune pesce rosso.
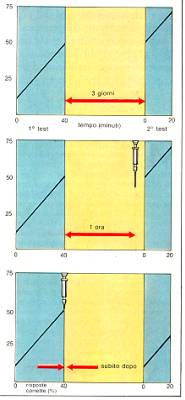
La somministrazione di un antibiotico a largo spettro come la
puromicina -che blocca la sintesi proteica e quindi la formazione di
nuove sinpasi- intreferisce negativamente col consolidamento di
un'esperienza. Quando la somministrazione della puromicina avviene
un'ora dopo l'esperienza, essa non è più efficace in
quanto la memoria a breve termine è già stata
consolidata grazie alla formazione di nuove sinpasi.
L’imprinting come modello di
memoria.

I pulcini si imprintano a uno stimolo
intermittente, più “dosabile” rispetto a uno stimolo
naturale.
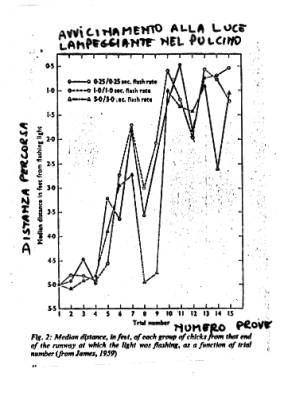
La curva dell'imprinting
è simile a quella dell'apprendimento
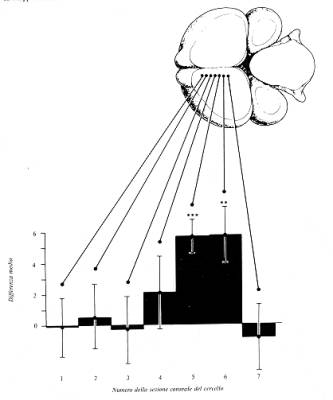
Nell’archistriato dei pulcini
imprintati si verifica un aumento della sintesi proteica, simile a
quello che caratterizza altre forme di esperienza.
La modulazione della
memoria
Il consolidamento viene moddulato da una
serie di fattori, ad esempio l'emozione. Alcuni mediatori nervosi o
modulatori come le endorfine intervengono in questo
processo.
1975 Gold e van Buskirk:
adrenalina post-trial ha effetti tempo e
dose dipendenti.
Behavioral Biology 13, 145,
1975.
Gli esperimenti su Aplysia
californica
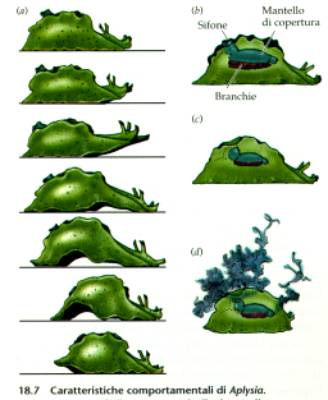
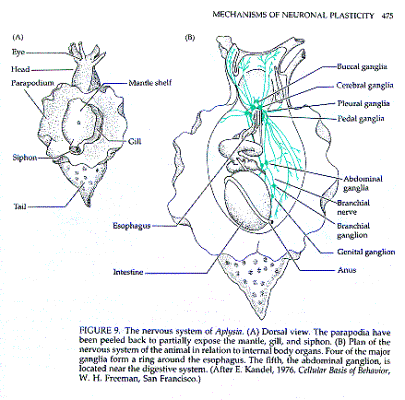
Lo studio dell'assuefazione dell'aplisia
ha fornito importanti notizie sul processo di memorizzazione.
L'aplisia si abitua a uno stimolo ripetuto, un sottile getto d'acqua:
l'abituazione è evidente in quanto il riflesso di retrazione
della branchia diminuisce col tempo. Ciò si accompagna a
modifiche neurochimiche a livello delle sinapsi.
E. Kandel Cellular bases of behavior, in
E.R. Kandel e J.H. Schwartz, Principles of neural science, pp.
396-408, Elsevier, New York, 1985
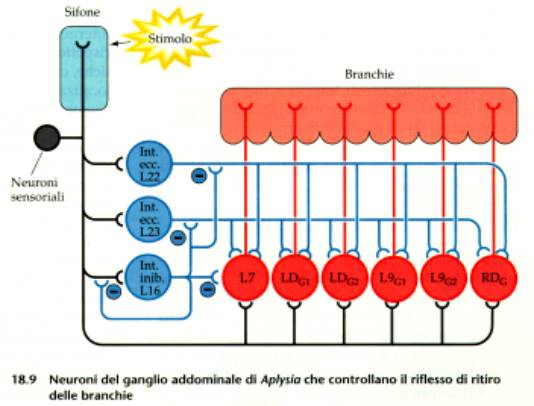
Tra i neuroni sensoriali e quelli motori
dell’Aplysia sono interposti interneuroni facilitanti o
inibitori.
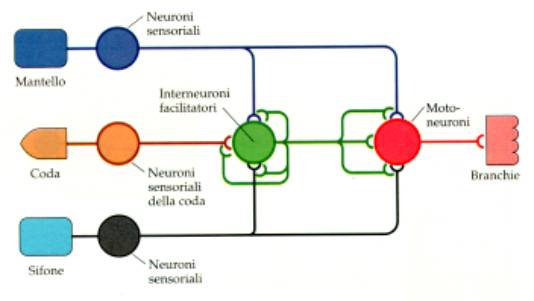
Nel
riflesso di retrazione della
branca e del sifone di Aplysia
possono intervenire, nel corso dell’assuefazione o della
sensibilizzazione, interneuroni che emettono sinapsi che esercitano
un effetto inibente o facilitante. Questi cambiamenti sinaptici sono
alla base delle forme più semplici di
esperienza.
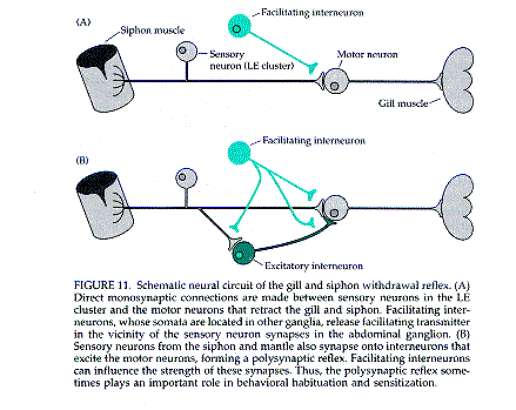
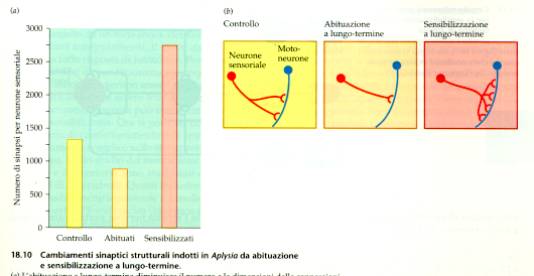
Modifiche sinpatiche nel corso
dell’abituazione e sensibilizzazione a lungo
termine.
Potenziamento a lungo termine o
“LTP”
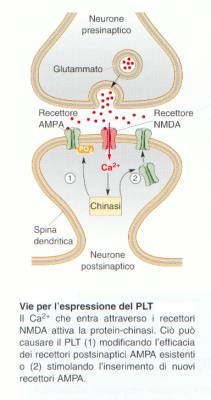
Il calcio che entra attraverso i
recettori NMDA attiva le protein-chinasi. Ciò può
causare l’LTP (1) modificando l’efficacia dei recettori
postsinaptici AMPA esistenti (2) o stimolando l’espressione di
nuovi recettori.

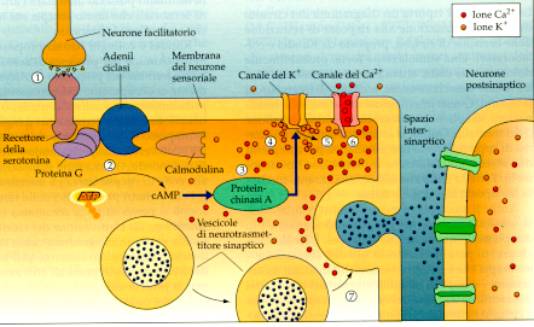
La cascata di eventi indotta
dall’attivazione di un neurone facilitatorio
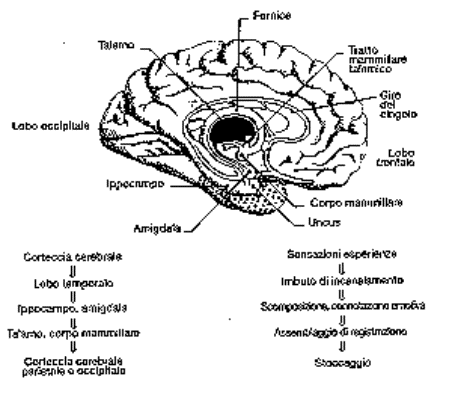
Ippocampo e corteccia
temporale
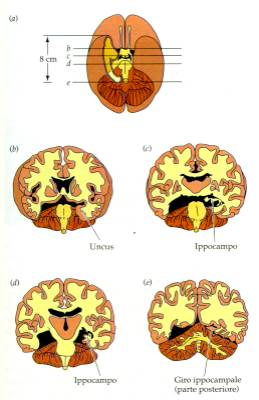
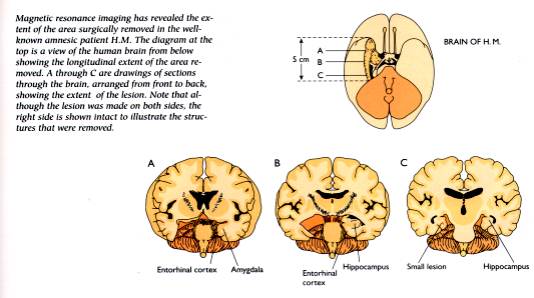
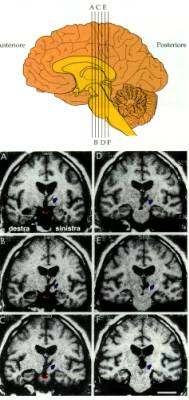
Un celebre caso clinico descritto da Milner e Scoville, il caso
del paziente HM ha chiarito come le lesioni dell'ippocampo e della
corteccia inferotemporale inducano gravi forme di amnesia.
Test per lo studio del comportamento
spaziale in rapporto al ruolo dell’ippocampo
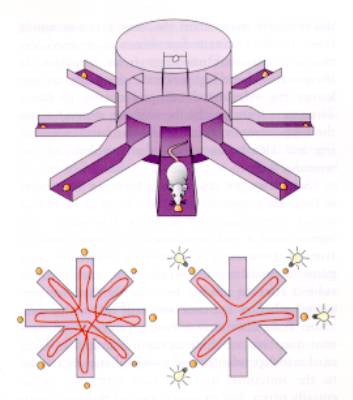
Labirinto radiale di
Olton
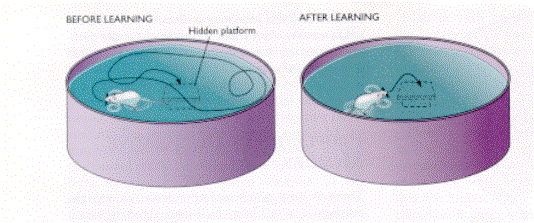
Water maze di
Morris
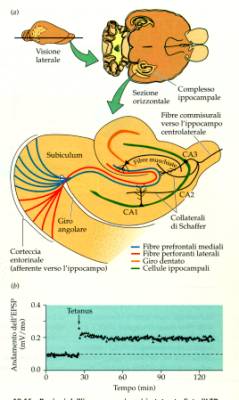
Circuiti
dell’ippocampo
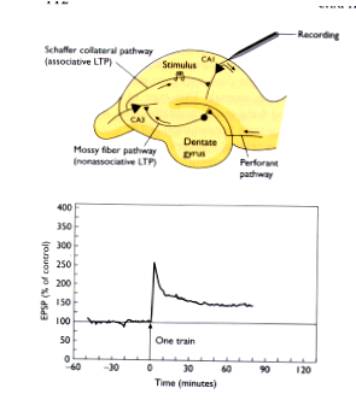
La stimolazione ripetuta di CA3
induce LTP in CA1
Ippocampo e memoria
umana.
Lesioni dell’ippocampo in H.M., un
celebre caso di amnesia studiato da B. Milner
W.B. Scoville e B. Milner, Loss of recent
memory after bilateral hippocampal lesions. Journal of Neurology,
Neurosurgery and Psychiatry, 20, 11-21, 1965
Brenda Milner e il MQ (Memory
Quotient):
si parla di amnesia quando una persona
dimostra un MQ più basso di 15 punti dell’IQ: nel caso di
HM, che aveva un quoziente di intelligenza medio, la differenza tra
l’IQ e il MQ era di ben 47 punti.
Lo striato ventrale e la
memoria
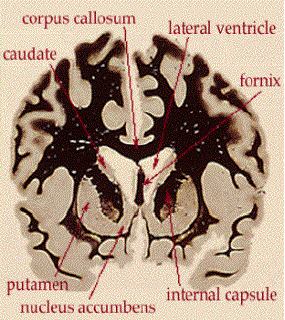
Posizione dello striato
ventrale
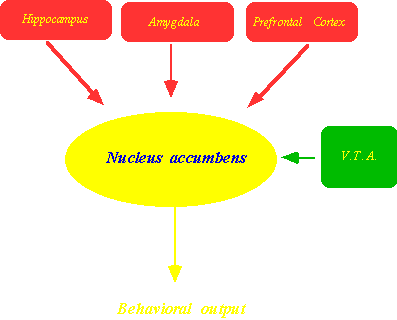
Rappresentazione schematica
degli input e output del nucleo accumbens
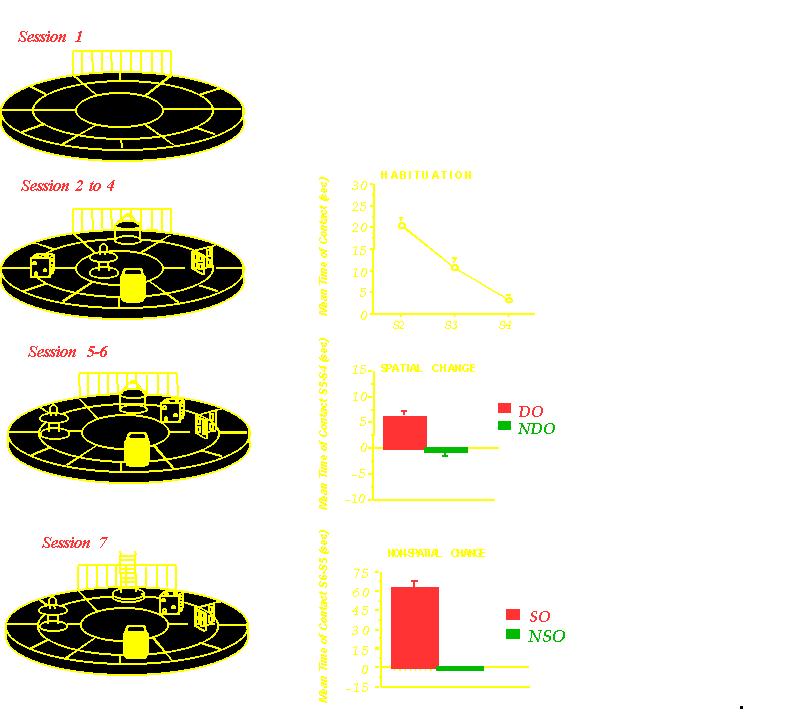
In questa arena si testano: Le reazioni
spaziali degli animali nei confronti degli oggetti spostati (DO) Le
risposte a oggetti nuovi Gli animali si abituano all’ambiente ma
reagiscono ai cambiamenti spaziali e non spaziali. Il loro
comportamento comporta memorie non rinforzate.
•L’accumbens è
coinvolto nel processamento dell’informazione spaziale, anche in
assenza di rinforzi.
•I recettori AMPA e NMDA (bloccati
con diversi antagonisti) modulano differenti aspetti del
processamento dell’informazione spaziale.
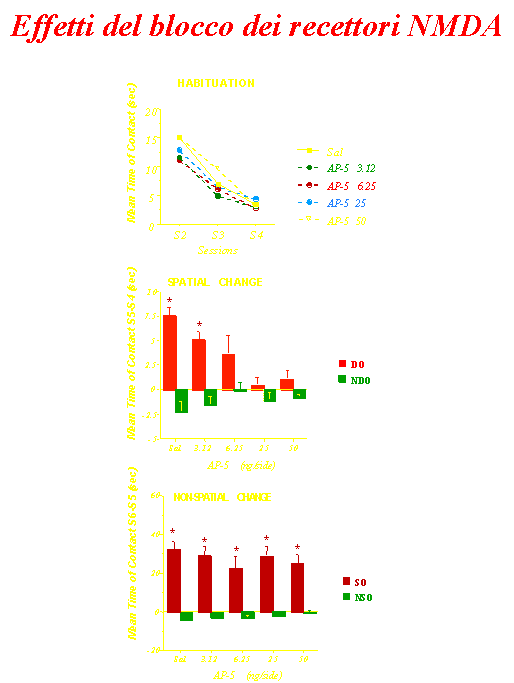
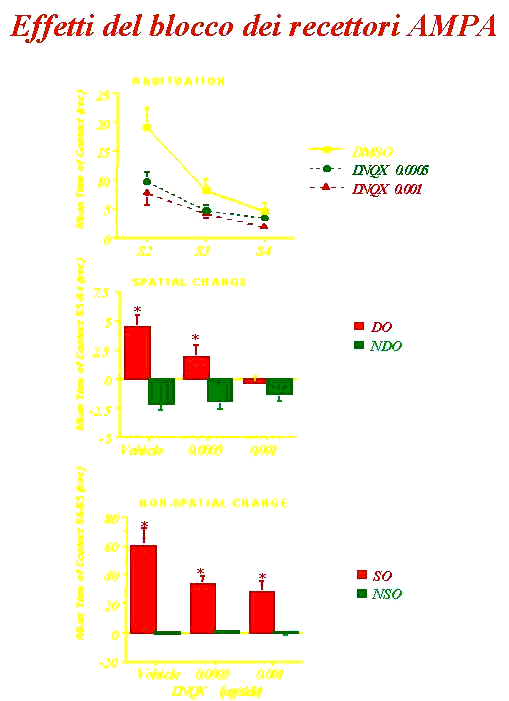
Lo striato ventrale è
un’interfaccia tra le funzioni cognitive e la loro espressione
comportamentale e gioca un ruolo critico nelle memorie e
comportamenti ricorrenti.
Corteccia cerebrale e
memoria
I ricordi di tutta la vita sono sepolti
nel cervello?
Marguerite Yourcenar:Quoi?
L’eternité.
“A lungo ho creduto di avere pochi
ricordi d’infanzia; intendo quelli anteriori al settimo anno. Ma
mi sbagliavo: penso piuttosto di non aver dato l’occasione
finora, ai ricordi, di risalire fino a me. Riesaminando i miei ultimi
anni al Mont Noir, alcuni almeno ritornano poco a poco visibili, come
fanno gli oggetti di una stanza dalla persiane chiuse nella quale non
ci si avventura da molto tempo.
Rivedo soprattutto piante e animali, su
un piano secondario rivedo alcuni giocattoli, giochi e riti che
avvenivano intorno a me, e più vagamente, come nello sfondo,
alcune persone”
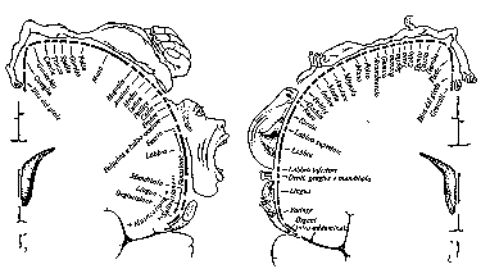
W. Penfield e le “mappe della
memoria”
(The mistery of the Brain,
1986).
Punto 5: “Solo un piccolo barlume
di un senso di familiarità e l’impressione che sapessi
tutto ciò che stava per accadere nel prossimo
futuro”.
Punto 12: “Sì, udivo voci
lungo il fiume da qualche parte: una voce di uomo e una voce di
donna… Penso che vedevo il fiume”.
Punto 17c: “Oh, avevo lo stesso
ricordo, molto, molto familiare, in un ufficio da qualche parte.
Riuscivo a vedere le scrivanie. Io ero là, e qualcuno mi stava
chiamando…”
Punto 18a: (stimolazione senza
preavviso): “Avevo un piccolo ricordo –una scena in un
gioco- stavano parlando e io potevo
vedere…”
Il numero degli stimoli in grado di
innescare una memoria diminuisce col tempo.
(Marigold Linton, 1986).
Per quindici anni ha annotato, giorno
dopo giorno, la descrizione di almeno un paio di eventi di rilievo: a
distanze varie, mesi o anni, ha notato che inizialmente i ricordi
sono vivi e non sono necessari molti suggerimenti (cue) per
rievocarli, ma man mano che il tempo trascorre il numero di
cue in grado di far ritornare alla mente quell’esperienza
si assottiglia sempre più.
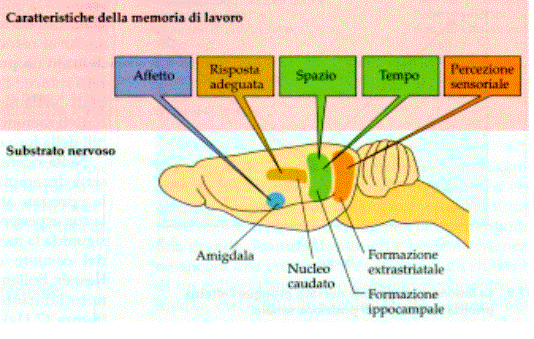
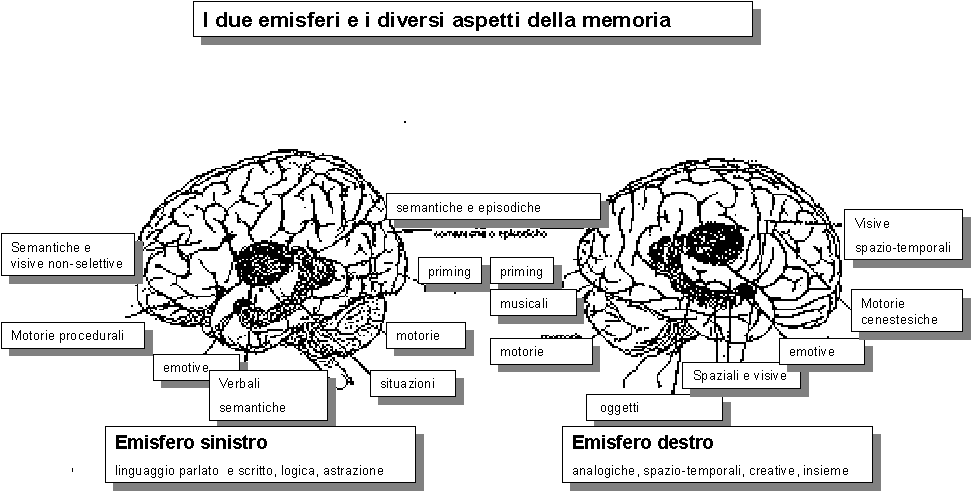
Qual è il codice dei neuroni? Il
codice della memoria?

Roger
Penrose: La memoria come fenomeno
quantico
Penrose, R. The Emperor's New Mind. New
York, Oxford Univ. Press, 1989
Il ri-consolidamento
1994 E. Loftus: La memoria incorpora
(falsi) dettagli di un evento che viene rievocato da
altri.
Eyewitness testimony. Cambridge, MA:
Harvard University Press.
Ri-consolidamento
1997 Roullet e
Sara: Consolidation of Memory
After its Reactivation: Involvement of b Noradrenergic Receptors in
the Late Phase J. Neural Plasticity, 6, 321, 1997.
2000 Nader, Schafe e
LeDoux: Fear memories require
protein synthesis in the amygdala for reconsolidation after
retrieval" Nature, 406, 2000.
2002 LeDoux e Nader:
Cellular and systems
reconsolidation in the hippocampus.
Neuron 36:527-38,2002.
Ri-consolidamento
Il blocco della sintesi proteica blocca
il consolidamento e non ha effetto prima del test. Il blocco della
sintesi proteica o una lesione dell’ippocampo induce un blocco
della memoria nell’animale riesposto all’esperienza
relativa al fear-conditioning.
Il ri-consolidamento come mezzo per
integrare i nuovi apprenimenti nelle esperienze
precedenti